
Artículo publicado en nuestra sección Hábitat COAMBA de la revista OTWO nº 50 del mes de septiembre de 2023. Desde la Página Nº 54 a la 67.
Autora: María José Romero Pérez. Ambientóloga. Alumna interna IBYDA (Instituto de biotecnología y desarrollo azul). Trabajo Fin de Grado en Ciencias Ambientales. Universidad de Málaga.
Los ecosistemas costeros son sistemas de transición entre el ambiente marino y el terrestre. Son de gran importancia, ya que son ambientes únicos por su constante dinamismo controlado por procesos químicos, físicos y biológicos. Todos estos procesos y ambientes cambian profundamente a escala geográfica, en el tiempo y en su duración haciendo que estos ecosistemas sean muy productivos, pero a la vez muy vulnerables (Botello et al., 2011). Dentro de estos ecosistemas, las macroalgas constituyen la mayor parte de la biomasa, alcanzando el 40% de la producción primaria, junto a las fanerógamas marinas. Las macroalgas son un grupo diverso filogenéticamente, constituido por organismos fotosintéticos en los que se incluye las algas marinas verdes, rojas y pardas.

En el intermareal, la distribución de las comunidades está controlada generalmente por factores físicos en las zonas más altas como son la inmersión, emersión, irradiancia, temperatura y disponibilidad de nutrientes, y por factores biológicos en las zonas bajas (Lewis, 1964; Sánchez de Pedro et al., 2013). La disponibilidad de nutrientes es un factor importante en cuanto a la distribución de las comunidades, por ejemplo, las especies que conviven en el mismo hábitat pueden competir por luz, sustrato y nutrientes (Lobban y Harrison, 1994). En este sentido, las macroalgas poseen distintos mecanismos para incorporar los nutrientes y por ello algunas especies son capaces de sobrevivir cuando escasean los nutrientes, gracias a sus altas tasas de absorción a bajas concentraciones y alta capacidad de crecimiento aprovechando los pulsos eventuales de nutrientes (Pedersen y Borum, 1997; Rosenberg et al., 1984; Sánchez de Pedro et al., 2013), mientras que otras especies de lenta absorción suplen esta desventaja con una alta capacidad de almacenamiento (Campbell, 1999; Ryther et al., 1981; Sánchez de Pedro et al., 2013; Young et al., 2009).
De acuerdo con esto, los cambios de las concentraciones de nutrientes pueden llevar a unas especies a dominar las comunidades y otras a desaparecer o reducir su presencia considerablemente, provocando una simplificación de la complejidad de las comunidades (Arévalo et al., 2007). Según describe Grime (1977) las estrategias que poseen las plantas ante factores estresantes se pueden clasificar en tres grupos: (1) Las especies competitivas, que ocupan zonas de bajo estrés y fluctuación, (2) Las tolerantes que son capaces de crecer bajo condiciones inestables y (3) las oportunistas, que son aquellas capaces de aprovechar los recursos al máximo en condiciones de contaminación. Esto nos puede dar una idea del estado de esas aguas según las especies que encontramos y su abundancia.
La gran riqueza y diversidad de los ambientes costeros ha llevado a una mayor concentración de actividades humanas en el litoral. Se estima que alrededor del 40% de la población mundial vive cerca o en la costa (IOC/UNESCO, IMO, FAO, UNDP., 2011). Las condiciones fluctuantes del intermareal y las presiones ligadas a las actividades antropogénicas hacen que sean aún más vulnerables a los efectos del cambio climático.

Uno de los principales cambios inducidos por el ser humano es el aumento de concentraciones de nutrientes en las costas por actividades como la construcción, el turismo, vertidos domésticos y la entrada difusa de fertilizantes nitrogenados y fosfatados, sobre todo de agricultura y ganadería.
La eutrofización hace que aumente la producción primaria, que en muchas ocasiones puede provocar hipereutrofización, con proliferaciones masivas de algas que generan condiciones de anoxia y por tanto un impacto negativo en el ecosistema (Campbell, 1999; Fonselius, 1978). Casos extremos hemos podido en el Mar Meno,r en Murcia, a causa de la agricultura intensiva y actividades humana donde se han llegado a tener valores de O2 por debajo de 1-2 mg/l llegando a provocar una mortalidad masiva de organismos marinos. Hering et al., (2010) señalaron la eutrofización como la causa más importante del estado de regresión de los ecosistemas costeros europeos, considerando sobre todo la reducción de vertidos de nutrientes como medida de restauración.
En los sistemas costeros la eutrofización causa un impacto a nivel global que afecta a los ciclos biogeoquímicos, produciendo cambios en las comunidades de algas bentónicas (Munda, 1993; Villares et al., 1998) y en comunidades zoo bentónicas (Norkko y Bonsdorff, 1996; Pihl et al., 1995; Villares et al., 1998). El estrés que puede generar la eutrofización de origen antrópico lleva a la simplificación de las comunidades y a una disminución de la riqueza específica. De este modo, se pueden reducir o desaparecer las especies llamadas ingenieras de ecosistemas (Fucales y Laminariales), que son formadoras de bosques y proporcionan grandes servicios ecosistémicos y ser sustituidas por especies de crecimiento rápido.; por ejemplo, E. Selaginoides (Linnaeus) Molinari & Guiry en el mar Mediterráneo puede sufrir una regresión en un eventual suceso de eutrofización; mientras que se favorece el crecimiento de
especies oportunistas como Ulva Rigida (C. Agardh ) (Rodríguez-Prieto & Polo, 1996).
Entre los nutrientes esenciales para las macroalgas se suelen incluir elementos como nitrógeno, fosforo, hierro y algunos metales como magnesio. El nitrógeno es uno de los elementos que puede ser limitante para el crecimiento de macroalgas y en algunos casos el fósforo (Lobban y Harrison, 1997).

También es importante conocer la ratio N:P limitante para una especie dada, ya que se ha podido comprobar que esto depende del género y la especie (Tilman et al., 1982). Por ejemplo, en las costas australianas se vio que para especies del género Ulva el factor limitante era el N y otras especies del género Cladophora estaban limitadas por la falta de fósforo (Valiela et al., 1987).
La Problemática de la eutrofización antrópica.
El problema de la eutrofización a causa de las actividades humanas en las costas es reconocido mundialmente como una amenaza importante para los ecosistemas costeros (Norkko y Bonsdorff, 1996; Valiela et al., 1997; Raffaelli et al., 1998; Sfriso et al., 2001). Una de las principales consecuencias sobre estos ecosistemas cuando aumenta la carga de nutrientes, es el cambio en las comunidades y el desplazamiento espacial de los productores primarios (Neto et al., 2012) como las macroalgas, afectando a las comunidades asociadas (macroinvertebrados, peces…) (Raffaeli et al., 1998) y a los servicios que generan al entorno (Jonge et al., 2000).
En este contexto de aumento de la eutrofización, las descargas de nutrientes de origen antrópico hacen que unas especies se vean más favorecidas que otras. La tasa de crecimiento, los requerimientos de nutrientes y la incorporación de éstos está directamente relacionado con la proporción superficie-área-volumen, por lo que las macroalgas filamentosas y foliosas crecen más rápido y precisan más nutrientes por unidad de biomasa y tiempo (Karez et al., 2004; Pedersen y Borum, 1997). En las costas rocosas eutrofizadas el crecimiento de algas filamentosas anuales aumenta, por lo que se puede retrasar la aparición de las especies perennes (Pedersen y Borum, 1996; Schramm, 1999). Esta sustitución por especies oportunistas es una señal de que hay una eutrofización creciente es esas aguas (Orfanidis et al., 2001).
El modelo de ciudad actual en Málaga es cada vez más incompatible con la sostenibilidad y con el mantenimiento de la calidad de las aguas costeras. El creciente aumento de las construcciones en el litoral no hace más que agravar la situación de deterioro de las costas. En concreto en la playa de la Araña, a pesar de no ser un lugar ocupado masivamente por el turismo, sí que encontramos otras infraestructuras que pueden afectar a los ecosistemas de esta playa. Principalmente la cementera, y posteriormente también incineradora de residuos, la cantera y la autovía.

La expansión que se está llevando a cabo en las costas malagueñas motivan el estudio de cómo el aumento de nutrientes de origen antrópico afectará a las tasas de incorporación de dos algas típicas de la costa: Ulva rigida, que además de tener una alta capacidad de producción, es una de las especies dominantes de la costa mediterránea (Figueroa et al., 2020), y Ericaria selaginoides, la cual se considera indicadora del buen estado de las aguas (Diez et al.,
1999), además de ser una especie que forma bosques marinos en el Mediterráneo.
Método de estudio
La incorporación de los nutrientes amonio, nitrato y fosfato se estimó siguiendo el modelo de la perturbación múltiple; método propuesto por Pedersen (1994) que combina la técnica de tubos múltiples y método de perturbación. Con esta técnica se mide la variación de los nutrientes en sucesivos intervalos de tiempo en medios de cultivos preparados con distintas concentraciones iniciales de sustrato. Este método ofrece información sobre todas las fases de
incorporación y la dependencia de las tasas de incorporación en el tiempo.
El experimento se llevó a cabo en un rango de concentraciones ambientalmente eutróficas, cuyas concentraciones iniciales fueron 100, 50, 25, 10 y 5 μM para amonio y nitrato, y 50, 25, 10 y 5 μM para fosfato.
Resultados
En este estudio las respuestas cinéticas de incorporación para ambas especies muestran claras diferencias según el nutriente y el tiempo. En general, Ulva rigida presentó mayores tasas de incorporación para los tres nutrientes. Constituyendo una ventaja en un contexto de eutrofización.
Los resultados obtenidos en este estudio apuntan a una ventaja de U. rigida sobre E. selaginoides por sus mayores tasas de incorporación para nitrato y amonio. Esto concuerda con las características descritas para las especies del género Ulva como oportunista (Mayorga, 2009) que además de su morfología externa, presentan altas tasas de crecimiento y una alta capacidad de incorporar y metabolizar nitrógeno inorgánico (fundamentalmente NO – y NH
+) (Cohen y Neori 1991; Littler y Littler, 1980; Rosenberg y Ramus 1984;). Mientras que E. selaginoides prospera en zonas poco afectadas por actividades antropogénicas y sin eutrofización (Bellan- Santini 1968; Belsher, 1979; Hoffmann et al., 1988; Ballesteros et al., 1998; Thibaut et al., 2005). Además, las macroalgas bentónicas como E. selaginoides pueden ayudar a identificar aguas perturbadas como bioindicadores (Ballesteros et al., 2007).
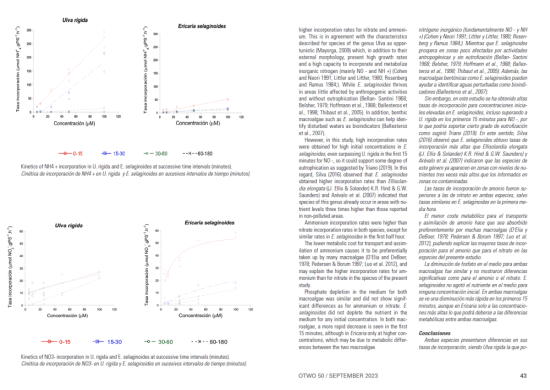
Sin embargo, en este estudio se ha obtenido altas tasas de incorporación para concentraciones iniciales elevadas en E. selaginoides, incluso superando a U. rigida en los primeros 15 minutos para NO -, por lo que podría soportar cierto grado de eutrofización como sugirió Triano (2019). En este sentido, Silva (2016) observó que E. selaginoides obtuvo tasas de incorporación más altas que Ellisolandia elongata ((J. Ellis & Solander) K.R. Hind & G.W. Saunders) y Arévalo et al. (2007) indicaron que las especies de este género ya aparecen en zonas con niveles de nutrientes tres veces más altos que los informados en zonas no contaminadas.
Las tasas de incorporación de amonio fueron superiores a las de nitrato en ambas especies, salvo tasas similares en E. selaginoides en la primera media hora.
El menor coste metabólico para el transporte y asimilación de amonio hace que sea absorbido preferentemente por muchas macroalgas (D’Elia y DeBoer, 1978; Pedersen & Borum 1997; Luo et al. 2012), pudiendo explicar las mayores tasas de incorporación para el amonio que para el nitrato en las especies del presente estudio.
La diminución de fosfato en el medio para ambas macroalgas fue similar y no mostraron diferencias significativas como para el amonio o el nitrato. E. selaginoides no agotó el nutriente en el medio para ninguna concentración inicial. En ambas macroalgas se ve una disminución más rápida en los primeros 15 minutos, aunque en Ericaria solo a las concentraciones más altas lo que podrá deberse a las diferencias metabólicas entre ambas macroalgas.
Conclusiones
Ambas especies presentaron diferencias en sus tasas de incorporación, siendo Ulva rigida la que posee una mayor Vmáx para el fosfato y amonio, y mayores tasas de incorporación de los tres nutrientes que E. selaginoides.
Ericaria selaginoides presentó menor afinidad por el amonio y nitrato que por el fosfato mientras que la afinidad por el fosfato fue similar en ambas especies. En líneas futuras de investigación se podría estudiar si E. selaginoides tiene mayor tolerancia a las aguas contaminadas y determinar en ambas especies la incorporación y acumulación de nutrientes y crecimiento a largo plazo, para corroborar los posibles cambios que se darían en la comunidad de macroalgas, considerando ambas especies.
En un contexto de eutrofización, U. rigida podría tener ventaja sobre E. selaginoides por la respuesta cinética y mayores tasas de incorporación de nitrógeno y fósforo obtenidas, pudiendo llegar a desplazar a esta última.
Finalmente cabe destacar la importancia del estado ecológico de las aguas, considerando la calidad biológica como la abundancia y presencia de especies de macroalgas bioindicadores, ya que éstas responden a los cambios del medio como la concentración de nutrientes o la hidromorfología (Neto et al., 2012). Por ello, hoy día la Directiva Marco Europea del agua (DMA2000/60/CE), exige a los estados miembros europeos evaluar el estado ecológico de las aguas, considerando así a las macroalgas como indicadoras.
Fuente: Revista OTWO nº 50 del mes de septiembre de 2023.
El colegio profesional no se hace cargo ni responsable de las opiniones o estudios publicados por parte de un colegiado o grupo de los mismos o fuentes externas.
COAMBA tiene una sección propia en la revista OTWO denominada Hábitat COAMBA. Si alguna persona Colegiada está interesada en escribir su propio artículo y que sea publicado en esta Sección, debe contactar con oficina@coamba.es para más información.
Animamos a nuestros/as colegiados/as a participar y a suscribirse para recibir publicaciones futuras de la revista OTWO de forma gratuita.